С днем рождения, профессор Гельфанд!
Поздравляем с днем рождения сооснователя «Диссернета» Михаила Гельфанда!
Как всегда, сегодня мы публикуем материал, не связанный непосредственно с «Диссернетом». И с радостью — опять, как всегда! — отмечаем, что нам есть, из чего выбрать. Отец-основатель сообщества верен идее просветительства и, несмотря на загруженность, успевает выступать перед самыми разными аудиториями и отвечать на множество вопросов.
Но не менее интересно наблюдать, как профессор Гельфанд вопросы задает. Поэтому пусть сегодня будет Гельфанд – отвечающий и Гельфанд – спрашивающий.
1. YouTube-канал ОСНОВА Борис Веденского
2. Фантастические гады и как их изучают. «Разговор за жизнь» с зоологом Еленой Темеревой
Naked Science, сентябрь 2022
Сухие, мокрые и полевые — на такие три типа биологи разделяют сами себя. Первые занимаются вычислениями, вторые ставят эксперименты в лабораториях, а третьи изучают биоразнообразие. Героиня этого «Разговора за жизнь» — профессор кафедры зоологии беспозвоночных биофака МГУ Елена Темерева — относится к последнему типу. Большую часть своей жизни она изучает беспозвоночных животных, разбирается в их строении, повадках и эволюции. О том, как устроена работа классического биолога в XXI веке, о своих открытиях и революциях в зоологии она рассказала в интервью Михаилу Гельфанду, биологу сухому. Эта беседа — для биологических гурманов. Naked Science публикует это интервью в рамках цикла «Разговоры за жизнь» — совместного медиапроекта Сколтеха, Российского научного фонда и Naked Science.
[Михаил Гельфанд]: Раньше вот думали, что биолог — это тот, кто с сачком ловит разных бабочек, высматривает тигров или, в крайнем случае, мучает мышек в клетке. Теперь «биолог» — человек в халате, с пробиркой или за микроскопом. Это действительно так, или «классическая» биология осталась?
[Елена Темерева]: Осталась, конечно, просто она обогатилась молекулярными методами.
[МГ]: Какими?
[ЕТ]: Если про мою работу — определяются последовательности генов разных червяков, строится филогенетическое древо (1), а потом мы его рассматриваем.
[МГ]: Почему именно червяков?
[ЕТ]: Потому что я червяками занимаюсь. Есть такое замечательное направление, филогеография (2), которым я планирую вплотную заняться в ближайшее время.
[МГ]: То есть классическая зоологическая таксономия стала строиться не только по анатомическим признакам, но и по молекулярным.
[ЕТ]: Именно так. Более того, сейчас является хорошим тоном, когда есть и качественно сделанная морфология, и молекулярка. Это не всегда возможно, конечно, потому что пробы разные бывают. У меня, например, большинство проб, которые мне привозят коллеги, либо формалиновые, либо в 70-процентном спирте; с ними дальше сложно работать. А так да, описание видов уже идет по-другому.
[МГ]: Что радикально поменялось в наших представлениях о таксономии с появлением молекулярных методов реконструкции филогенетических деревьев?
[ЕТ]: Много чего. Хотя бы такой простой пример: оказалось, что есть очень много криптических видов (3), которые мы раньше не умели различать ни на морфологическом, ни на анатомическом уровне, а после молекулярного анализа становится понятно, что есть два разных вида, и начинаешь копать, а что, собственно, разного. Должно же что-то быть? Морфология, анатомия, развитие… Может вовсе ничего не находиться, а может находиться в самом неожиданном месте. Например, оказывается, что у них личинки совсем разные, у двух якобы одинаковых видов.
[МГ]: Совсем разные или немножко разные?
[ЕТ]: Это как посмотреть: может быть, и немножко, а может быть, и сильно разные. Особенно это, конечно, важно для тех видов и вообще тех групп, у которых мало всяких морфологических признаков. Я занимаюсь форонидами (рис. 1), внешне они все абсолютно похожи друг на друга, определить их на глаз даже я не смогу — их нужно резать, делать срезы и 3D-реконструкции каких-то органов. Тут молекулярный анализ, конечно, очень помогает.

© Антон Чичвархин
[МГ]: Какая разница, сколько там видов?
[ЕТ]: Это биоразнообразие.
[МГ]: Если их нельзя различить, какое ж там разнообразие?
[ЕТ]: Всякое познание биоразнообразия имеет смысл, потому что разные виды живут в разных биотопах, и если говорить о форонидах, то это виды-эдификаторы (4) (рис. 2) — они часто образуют очень плотные поселения, могут покрывать дно многокилометровым ковром. Понятно, что они определяют состав сообщества. Разные виды форонид связаны с разными другими видами: одни живут всегда в ассоциации с какими-то аннелидами, с конкретным видом полихет, а другие, например, с моллюсками.

© Антон Чичвархин
[МГ]: Двойники, которые неразличимы, тоже образуют разные сообщества?
[ЕТ]: Этого я доподлинно не знаю, но думаю, что да.
[МГ]: То есть пока не было молекулярки, никому не приходило в голову обратить внимание, что есть скопления вроде как одинаковых форонид, которые при этом образуют ассоциации с совсем разными организмами, и подумать, что, может быть, это не случайность, а они-таки действительно разные, и начать смотреть морфологию.
[ЕТ]: Абсолютно так, да. Более того, если говорить про эти гидробиологические работы, просто сбор всего макробентоса конкретной акватории, то форонид вовсе не определяют, то есть форониды и всё, только до ранга типа. Некоторые, правда, и этого не могут, то есть пишут аннелиды, а оказываются — форониды.
[МГ]: Это у кого-то ручки кривые и глазки косые.
[ЕТ]: Все равно, тип — это вовсе не ранг вида.
[МГ]: Хорошо, это, так сказать, микротаксономия. Думали, что один вид, а оказалось, что пятнадцать. А глобальные изменения? У Линнея было легко: млекопитающие, птицы, гады, рыбы, насекомые и черви — и в червях все это радостно сидело.
[ЕТ]: На самом деле, молекулярка сделала просто невероятный прорыв в большой филогении, по крайней мере, филогении билатерий (5) и, на самом деле, вообще многоклеточных. Что я имею в виду? Кеннет Халанич с соавторами в 1995 году показали, что те самые форониды — родственники первичноротых животных, а не вторичноротых, как всегда считалось…
[МГ]: Для «чайников» нужно объяснить про первичноротых и вторичноротых.
[ЕТ]: Первичноротые — это, например, кольчатые черви и моллюски, а вторичноротые — это мы с вами, иглокожие и несчастные полухордовые, которых мало.
[МГ]: А насекомые?
[ЕТ]: Насекомые — это к Ecdysozoa (6), это следующий мой пункт. В 1997 году Агвинальдо с соавторами в журнале Nature написали, что нематоды — это родственники членистоногих. Была выделена группа Ecdysozoa, и сейчас вообще вся система билатерий не похожа на то, что было до 1995 года, потому что раньше было: паренхиматозные, первичнополостные, вторичнополостные, вот это всё (рис. 3). А оказалось вдруг, что нематоды и раки — это ближайшие родственники.

Рис 3а.
 и новые (б) представления о системе и филогении билатерально симметричных животных.1 — свободноживущие плоские черви (турбеллярии), 2 — паразитические плоские черви, 3 — немертины, 4 — круглые черви (нематоды), 5 — головохоботные черви, 6 — коловратки, 7 — кольчатые черви, 8 — моллюски, 9 — сипункулиды, 10 — хелицеровые, 11 — ракообразные, 12 — насекомые, 13 — форониды, 14 — мшанки, 15 — брахиоподы, 16 — полухордовые, 17 — иглокожие, 18 — хордовые. На схеме представлены далеко не все группы животных.Источник: Малахов В.В., Революция в зоологии. Природа. 2009. №3. С. 40-54.")
Рис 3б. Классические (а) и новые (б) представления о системе и филогении билатерально симметричных животных.
1 — свободноживущие плоские черви (турбеллярии), 2 — паразитические плоские черви, 3 — немертины, 4 — круглые черви (нематоды), 5 — головохоботные черви, 6 — коловратки, 7 — кольчатые черви, 8 — моллюски, 9 — сипункулиды, 10 — хелицеровые, 11 — ракообразные, 12 — насекомые, 13 — форониды, 14 — мшанки, 15 — брахиоподы, 16 — полухордовые, 17 — иглокожие, 18 — хордовые. На схеме представлены далеко не все группы животных.
Источник: Малахов В.В., Революция в зоологии. Природа. 2009. №3. С. 40-54.
[МГ]: Много крови пролилось, пока это оказалось.
[ЕТ]: Да, потому что это невозможно было принять: членистоногие оказались оторваны от аннелид (7), а эта группа Articulata была в каком-то лохматом году еще Кювье выделена… членистоногие выводились напрямую из аннелид. Так что большую филогению просто перетрясло от этих молекулярных штук, но в таком виде она теперь абсолютно устойчива. Есть Spiralia, есть Ecdysozoa, есть вторичноротые. Теперь мы так и живем, в этой парадигме.
[МГ]: Насколько эта парадигма устойчива? Не окажется еще через двадцать пять лет, что все было не так?
[ЕТ]: Я думаю, что не окажется, потому что она имеет и морфологические подтверждения, и молекулярные.
[МГ]: Откуда взялись морфологические подтверждения? Предыдущая таксономия была основана на морфологии, дальше пришли молекулярные люди, перерисовали все деревья, а морфологи такие говорят: «А, ну так тоже годится. Мы по вашим новым деревьям теперь правильные морфологические признаки найдем».
[ЕТ]: Я понимаю, к чему вы клоните. Эти экдисозойные войны шли долго. У нас на первом курсе биофака новая система начала читаться не так давно, всего лет десять назад. Потому что это невозможно было принять в голову. Но на самом деле, действительно, есть и морфологические общие черты, которые пропускали… Не то что пропускали… Которым, может быть, не придавали такого значения.
[МГ]: Имеет ли смысл вообще понятие типа? Вид — в первом приближении понятно — это те, кто друг с другом спариваются, а с другими не спариваются. А вот все остальные ранги таксонов имеют смысл или это некоторая попытка натянуть на непрерывное дерево какую-то дискретную иерархию?
[ЕТ]: Конечно, попытка. Это все измышления человеческого мозга, но как-то же нужно систематизировать окружающее.
[МГ]: Вы же оперируете этими понятиями. Вот вы говорите: тип. Но тип хордовые и тип, скажем, полихеты — одновременно ли они образовались, если сравнивать по времени самые первые разветвления внутри типов? Или хордовые моложе, чем полихеты? Но тогда почему мы и то, и другое называем типами?
[ЕТ]: Все в кембрии еще появились.
[МГ]: А до кембрия никого многоклеточного не было?
[ЕТ]: Была, конечно, вендская фауна. Все эти дикинсонии и прочие товарищи.

Фоссилизированный Dickinsonia сostata в породах эдиакарского периода.
/ © Verisimilus, English Wikipedia
[МГ]: И чьи потомки выжили?
[ЕТ]: Кто ж знает-то? Например, не так давно описаны из докембрия якобы лофофораты. Меня это интересует, потому что форониды — это лофофораты. И вот, строят какие-то очень странные реконструкции. Я не палеонтолог, поэтому мне сложно судить, что из этих находок вообще можно извлечь… Но реконструкции выглядят странно, и в таком виде, конечно, никакие лофофораты до нас не дожили. Но если следовать логике авторов, то это был некий предок, а дальше пошла радиация в кембрии.
[МГ]: Если я правильно понимаю, все нынешние таксоны уровня типа в каком-то смысле появились в кембрии как отдельные ветви дерева, да?
[ЕТ]: Либо не как отдельные ветки, может быть на уровне каких-то предковых форм.
[МГ]: Если мы выделяем эти предковые формы, значит они отделились от прочих, а потом пошла радиация.
[ЕТ]: Да, как-то так.
[МГ]: Первое ветвление на предков современных типов — это кембрийский взрыв. Правильно?
[ЕТ]: Да, видимо, так.
[МГ]: Стало быть, понятие таксона уровня типа имеет смысл, потому что это одна быстрая радиация, так же как и отряды млекопитающих имеют смысл, потому что они 70 миллионов лет назад начали все резко ветвиться.
[ЕТ]: Я просто никогда не рассматривала это через призму какой-то временной шкалы. Мне казалось, что тип — это животные, которые обладают уникальным планом строения.
[МГ]: Вы верите в парафилетические таксоны (8)?
[ЕТ]: А это вопрос веры? [Смеется.]
[МГ]: Хорошо, готовы ли вы в своей научной практике допустить использование парафилетических таксонов?
[ЕТ]: Думаю, что да.
[МГ]: Именно потому, что у них общая морфология? А то, что у них изнутри торчит длинная чужая ветка, так бог с ним? Вот классический пример с китами, которые не так давно оказались парнокопытными. Копыт, конечно, у них никаких нет, но они, тем не менее, молекулярно парнокопытные, железно, никуда не денешься. И последними решениями партии и правительства отряд китообразных упразднили.
[ЕТ]: Вот этого я не знала!
[МГ]: Всё, нет никаких китообразных и парнокопытных. То есть люди взяли и, наплевавши на морфологию, пинками установили монофилетический отряд китопарнокопытных ровно затем, чтобы не было парафилитического отряда. И это действительно вопрос веры.
[ЕТ]: Согласна, да. Хотя мне, конечно, как морфологу, сложно смотреть на китов как на парнокопытных.
[МГ]: А на птиц как на рептилий?
[ЕТ]: Проще, потому что это как-то из школы.
[МГ]: Змей, кстати, тоже отменили.
[ЕТ]: А они теперь кто?
[МГ]: Ящерицы.
[ЕТ]: Змеи — ящерицы. Хорошо хоть не парнокопытные.
[МГ]: Они, кажется, ближе к игуанам, хамелеонам и варанам, а потом уже сцинки и гекконы. В общем, змеи сидят глубоко внутри ящеричного дерева.
[ЕТ]: Понятно.
[МГ]: А что, кафедры беспозвоночных и позвоночных друг с другом научными новостями не обмениваются? Потому что, казалось бы, со змеями и с китами — это большие события.
[ЕТ]: Про змей я не была в теме… Вообще, это большая проблема. Мы часто сидим на кафедре и не знаем, что в соседней комнате делается. Поэтому на кафедре мы организовали семинар. Оказалось ужасно полезно, в первую очередь для нас самих. Была попытка сделать межкафедральные семинары на уровне ученого совета, и у нас они были одно время на регулярной основе, но потом все это затухло, потому что людям не до того, надо же делать доклад, к нему готовиться долго. Это очень жаль, потому что, например, ты слушаешь докладчика, какого-нибудь микробиолога: «Боже мой, я же видела бактерий… надо вместе сделать какую-нибудь интересную работу по бактериому каких-нибудь…»
[МГ]: Конечно, на червяках куча бактерий сидит.
[ЕТ]: И на них, и в них, и в трубках и где угодно.
[МГ]: Много народу ходит на ваши семинары по зоологии?
[ЕТ]: Очень зависит от темы. Леонид Русин делал доклад, как новые технологии, single cell анализ (9), помогают понять происхождение многоклеточных. Очень здоровский, хорошо построенный, понятный, потому что для махрового зоолога все эти single cell — это темный лес.
[МГ]: Да ладно, чего там сложного?
[ЕТ]: Ну, не знаю. В общем, тема интересная, пришло много народу.
[МГ]: А на какие семинары мало народу ходит?
[ЕТ]: На специальные. У нас был семинар про развитие брахиопод, это очень интересная тема, мы с Татьяной Кузьминой делали эту работу. Мне показалось, что мы сотворили машину времени, потому что мы реконструировали жизненные циклы вымерших брахиопод — их же гораздо больше, чем современных, и как они жили, что делали, как размножались, об этом ничего неизвестно. Но это интересно, может быть, палеонтологам и современным лофофоратчикам, а большинству народа, наверное, нет, поэтому на этот семинар мало людей пришло.
[МГ]: Студенты ходят?
[ЕТ]: Студенты нечасто, в основном аспиранты ходят. Некоторые заинтересованные студенты тоже, какие-нибудь олимпиадники. Поначалу им кажется, что все надо везде узнать, а потом этот интерес как-то очень быстро пропадает.
[МГ]: Возвращаясь к тому, о чем мы начинали, про модную биологию и немодную, на какие кафедры идут биологические олимпиадники?
[ЕТ]: Очень по-разному. Я в прошлом году вела в Звенигороде (10) группу, она была из олимпиадников, и был такой очень выдающийся молодой человек, который сказал: «Конечно, я пойду на молекулярную биологию, потому что молекулярная биология вообще царица биологии и все такое». Я ему объяснила, что молекулярная биология, конечно, да, но методы применяются везде, и у нас тоже, а знать разнообразие полезно. В итоге он пошел на нашу кафедру.
[МГ]: То, что вам понравилось про единичные клетки, это все-таки не совсем молекулярная филогенетика, это уже другая наука.
[ЕТ]: Да, какая-то другая.
[МГ]: Может быть, ее всем выучить в обязательном порядке?
[ЕТ]: Я думаю, что всем не нужно заниматься всем, потому что это бесперспективно. Хорошо быть специалистом, а не всё по верхам знать.
[МГ]: Часто самое интересное, это когда человек хорошо знает классическую биологию и видит прикольные объекты, на которые у других руки не дошли посмотреть. Но при этом одного понимания зоологической экзотики недостаточно, потому что нужно еще понимать, что с ними интересного можно сделать. И если не знать, что есть, например, анализ транскриптомов, то просто в голову не придет этим попользоваться.
[ЕТ]: Естественно, нужно иметь представление о наборе методов. Но так, чтобы быть специалистом…
[МГ]: Уметь считать необязательно, да. Найдется тот, кто умеет считать, это не проблема.
[ЕТ]: На это вся надежда.
[МГ]: Желающих посчитать обычно больше, чем тех, кто понимает, что именно надо посчитать.
[ЕТ]: [Cмеётся.] Абсолютно согласна. У наших зарубежных коллег есть, мне кажется, не то что проблема, я не готова их как-то судить…
[МГ]: Зоологии нету хорошей.
[ЕТ]: Да, и это ведет к неким проблемам, потому что у них есть методы, очень классное оборудование, они в это оборудование засовывают все что ни попадя… Вот экспрессия генов: засунули туда форонид. Зачем они это сделали, совершенно непонятно. Какой результат получился, тоже непонятно. Описание не то что неправильное, а просто перевернутое с ног на голову. В общем, это только расстраивает. Я такие работы даже иногда не знаю, как цитировать, потому что нашли они, что какой-то ген экспрессируется вот там-то, а дальше-то что? Что из этого следует? Я бы хорошую работу сделала по экспрессии генов с форонидами и брахиоподами.
[МГ]: Если бы у вас был хороший аспирант, который умеет ставить транскриптомный эксперимент, потом посчитать, и, скажем, какой-нибудь грант, чтобы это все сделать — что бы вы сделали с форонидами и брахиоподами?

[ЕТ]: В первую очередь я бы выяснила происхождение их плана строения. Нужно сделать экспрессию на дорсальную и вентральную сторону (11). Есть же какие-то гены, которые маркируют эти стороны. Потому что есть мнение, что группы эти родственные, более того, некоторые молекулярные филогенетики помещают форонид просто внутрь брахиопод, не то что это отдельный тип…
[МГ]: Вот, а вы мне только что говорили, что Господь Бог как создал типы в кембрии, так они все радостно и ползают, а теперь оказывается, что форониды сидят внутри брахиопод.
[ЕТ]: Да, согласно товарищу Кохину. Это такой исследователь из Британии. Я не знаю, откуда у него эта идея. Но он молекулярщик, и у него вообще на эту тему странные представления, как раз модные.
У меня в 2015 году в PLoS One вышла статья, где я морфологически попыталась показать монофилию лофофорат, что форониды, брахиоподы и мшанки — это монофилетичная группа, а он мне написал письмо: «Куда вы, морфологи, лезете в построение филогении. Это мы должны делать». Вежливо так, но с напором, что морфология не в состоянии это всё решить, и оставьте это нам, молекулярным филогенетикам. У него, соответственно, есть работа, в которой написано, что форониды — это брахиоподы без раковины. При этом план строения у этих двух групп абсолютно разный.
[МГ]: Погодите, у нематод и насекомых тоже план строения неодинаковый.
[ЕТ]: Слава богу.
[МГ]: А все-таки согласились с молекулярными деревьями, деваться некуда было, приперло.
[ЕТ]: Да, но это все-таки две разные группы. Нематоды — тип. Вроде пока этого никто не отменял. А насекомые пусть будут артроподами. А форониды не должны быть в брахиоподах. они сложились на спинную сторону, а брахиоподы на брюшную. Я думаю, что это принципиальная разница. Мы видим признаки этого складывания в личиночном развитии брахиопод, но при этом есть разные мнения: кто-то видит, а кто-то не видит; кто-то говорит, что это вообще не складывание, а эти личинки немножко подвернулись и всё. А вот экспрессия нам бы помогла выявить гены, которые маркируют дорсальные и вентральные стороны на личиночных стадиях, на ювенильных (12), то есть в процессе метаморфоза, что очень важно, а уже у взрослых более или менее понятно.
[МГ]: Вы думаете, что экспрессия этих генов-маркеров между типами сохраняется?
[ЕТ]: Я думаю, что да, почему бы нет?
[МГ]: Я тоже не знаю. Есть дорсально-вентральная ось у насекомых, и есть у позвоночных. И те, и другие ползают на животе. Одни и те же гены отвечают за эту дифференцировку?
[ЕТ]: Я не знаю, так я не углублялась. Эта идея возникла, когда Русин пришел на семинар про брахиопод и сказал: «Чего вы вообще, морфологи, мучаетесь со своими личинками?»
[МГ]: Сначала надо посмотреть кого-нибудь известного и понять, сохраняются на этих основные регуляторы экспрессии или нет. А то может оказаться, что на таких расстояниях все развитие другое, и вы все равно ничего не увидите. В общем, мораль такая, что если есть полные геномы тех зверей, которых вы умеете растить, то это все можно сделать. Другое дело, как объяснить, что это важное и полезное импортозамещение.
[ЕТ]: Да, теперь именно так все надо объяснять. Я хотела посмотреть регенерацию, это сейчас тоже такая модная импортозамещающая тема. Форониды очень легко регенерируют. Могут просто голову отрастить: во-первых, им откусывают регулярно рыбы, раки, моллюски, а во-вторых, они просто после оплодотворения могут сами голову терять в буквальном смысле слова и быстро ее восстанавливают.
[МГ]: По нынешним временам это должно легко продаваться.
[ЕТ]: Да-да. Мы хотим посмотреть сначала на анатомическом, потом на клеточном уровне всякую пролиферацию, а потом сделать протеомный анализ с коллегами из Испании.
[МГ]: Всякое модное еvo-devo (13) до вас дотекло?
[ЕТ]: До нас дотекло, но вот конкретно до меня не очень. А на кафедре у нас некоторые работают этими методами, но, в общем, с разным успехом. Нужно иметь лабораторию, где это можно было бы делать нормальным образом. Мы сейчас как раз пытаемся что-то такое сделать. Но теперь это уже сложно будет.
[МГ]: Представим себе, что мы разговариваем не сейчас, а полгода назад, и представим себе, что, вы руководитель большого проекта, поэтому у вас денег немерено. Что бы вы стали делать из того, что вам интересно не для человечества, а для себя?
[ЕТ]: Ого! [Смеется.] Ничего себе. Не для человечества, а для себя? Не знаю, я как-то растерялась.
[МГ]: Вы себе никогда не задавали такого вопроса, чего бы хотелось реально?
[ЕТ]: Нет, ну хотелось бы, конечно… Я бы купила всякое чудесное оборудование.
[МГ]: А задачи-то какие?
[ЕТ]: Задачи… Задачи, наверное, изучение морфологии…
[МГ]: Для изучения морфологии никакие безумные приборы не нужны. В крайнем случае, хорошие микроскопы.
[ЕТ]: Хорошие микроскопы дорого стоят.
[МГ]: Хорошо. Вот вы закупились хорошими микроскопами и наразглядывались морфологию. Что дальше?
[ЕТ]: Дальше таксономия на основании морфологии и, наверное, молекулы.
[МГ]: Замечательно. Насеквенировали геномов, нарисовали филогенетических деревьев, натыкали на эти деревья морфологические изменения, чтобы понять, какой содержательный смысл в этих ветках. А как же с молекулярными деревьями, которые форонид посадили внутрь брахиопод? А вы говорите, что не верите в эти молекулярные деревья, потому что морфология совсем другая. Как вы на основании морфологических соображений оцениваете молекулярные деревья, как правдоподобные или как неправдоподобные?
[ЕТ]: Ну вот такие. Например, щетинкочелюстные, это морские стрелки, очень странный тип, который поносило по древу билатерий, где он только не был. Его сначала морфологи с вторичноротыми сближали, потому что у них такая анатомия, три целома (14), радиальное дробление яйца (15). А молекулярщики что только с ними ни делали! И экдизозоями они были, и в лофотрохозоа они были. А сейчас вышло несколько статей в Nature, и их угнездили в Gnathifera (рис. 5) — это такая группа, у которых есть челюсти. Туда относятся очень странные существа: коловратки вместе со скребнями (16), микрогнатозои (17) и гнатостомулиды (18). У них есть очень специфический челюстной аппарат, мастокс, то есть челюсти внутри глотки (рис. 6), которые могут выбрасываться. И вот эти челюсти сравниваются молекулярными филогенетиками со щетинками щетинкочелюстных, у которых есть ловчий аппарат, состоящий из пар щетинок на голове.
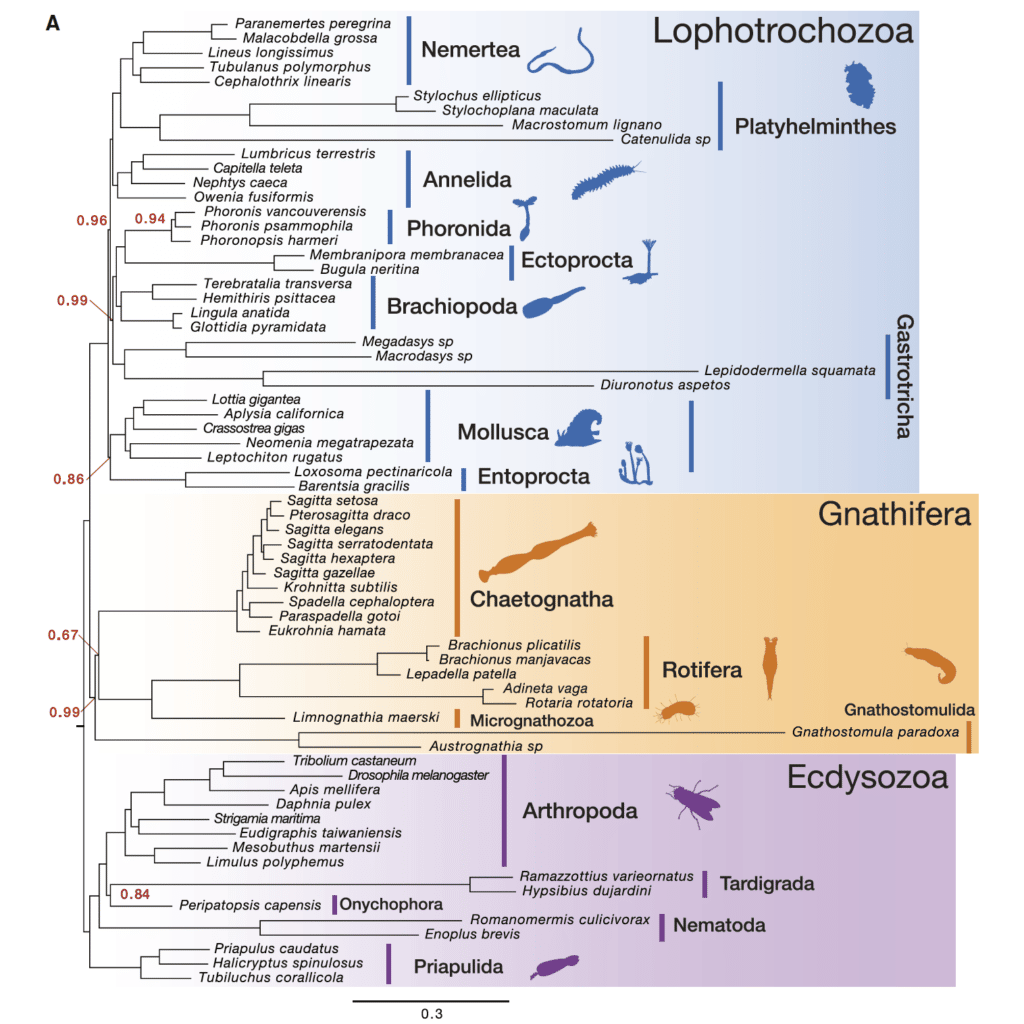
Источник: Marletaz et al., A New Spiralian Phylogeny Places the Enigmatic Arrow Worms among Gnathiferans. Current Biology. 2019. №29. P. 312-318.
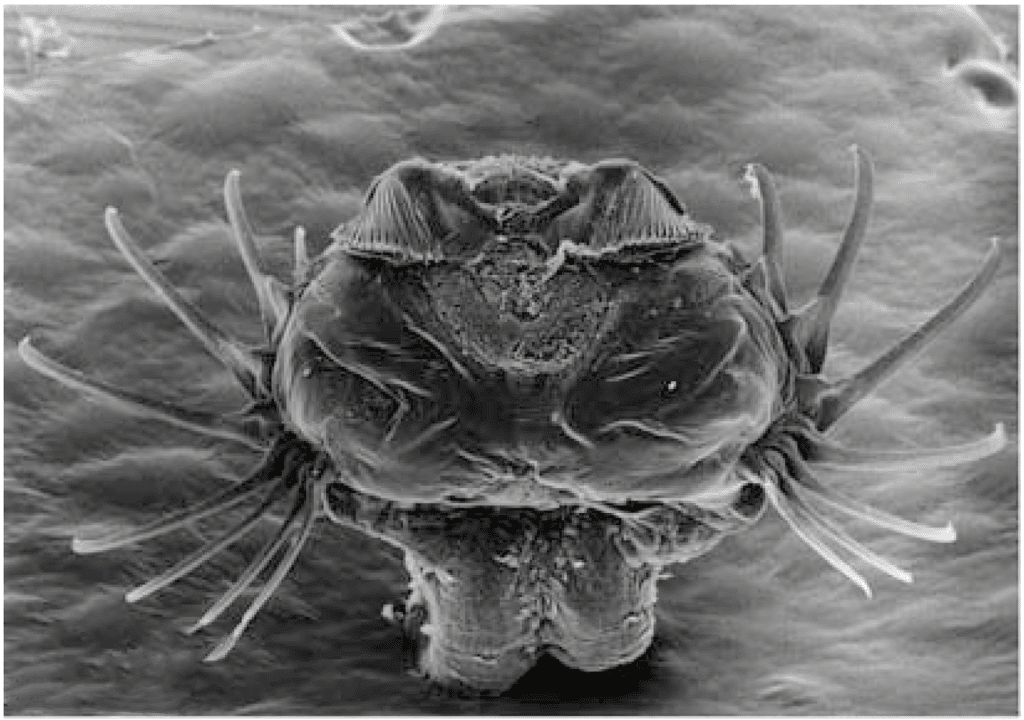
Но вот вопрос: во-первых, ладно, это всё эктодермальные структуры (19), что во рту, что на голове, но как-то странно: челюсть, выползшая на голову, — как они себе это представляют? Более принципиально — это то, что строение этих челюстей совершенно другое (вот зачем нам нужна вся эта морфология, в том числе на электронном уровне, а не только на световом), потому что у щетинкочелюстных щетинка — это как зуб: внутри нее есть пульпа, состоящая из клеток, которые являются продолжением эпидермальных клеток, а у нормальных гнатифер это просто кутикула, в которую не проходят никакие клетки, ни даже отростки клеток, максимум там могут быть микроворсинки. Я не считаю, что люди, которые сделали щетинкочелюстных гнатиферами, совершенно неправы, но это тот самый случай, когда это нужно потестить именно морфологическими методами.
Или мшанки несчастные… их тоже, бедных, поносило по дереву. В Ecdysozoa их никогда не относили, но среди Lophotrochozoa они тоже побывали, и где-то в основании древа. Они, наверное, все-таки лофофораты.
[МГ]: Когда кого-то так носит по дереву, это означает, что веточка, на которой он сидит, очень короткая и поэтому ненадежная.
[ЕТ]: Да.
[МГ]: Это означает, что по-хорошему никакого бинарного дерева нет, а есть просто большие узлы, из которых растет сразу несколько веток, которые поэтому толком и не разрешаются. А если там неразрешенный узел, из которого растут все трое, то не окажется ли так, что всегда найдутся общие морфологические признаки, которые любых двух отделят от третьего просто потому что они, на самом деле, растут из одного узла?
[ЕТ]: Тут сложно. Если говорить конкретно про мшанок, то их традиционно объединяли с животными, которые просто по размерам маленькие. Мшанки образуют колонии, но сами индивидуумы, зооиды, они мелкие. Во-вторых, у них есть какие-то ресничные аппараты, поэтому их объединяли с колокольчиковыми и с циклиофорами.
[МГ]: Тоже мне big deal, ресничный аппарат — да хоть у сувоек он тоже есть.
[ЕТ]: У сувоек он по-другому устроен, это одноклеточные существа. Считается, что у циклиофор сначала были щупальца, потом они уменьшались, а поскольку циклиофоры вообще очень маленькие, то от щупалец остались только реснички.
[МГ]: От каждого щупальца осталась одна клетка?
[ЕТ]: Ну например. Или пучок клеток.
И вот все эти малыши сгоняются в одну группу, которую Андреас Хайноль с соавторами в 2009 году назвали Polyzoa.
[МГ]: Это примерно как черви у Линнея.
[ЕТ]: Наверное. Мшанки там и оставались довольно долго, но на самом деле у них есть лофофор. Это очень специфический орган, и у разных животных он очень сходно организован. Мы сейчас как раз пишем статью в специальный номер Journal of Morphology, который будет называться Homology, и как раз пытаемся эту мысль о гомологии лофофора изложить. А у колокольчиковых есть щупальца, но это не лофофор, потому что у них и рот, и анус находятся внутри венчика, а это противоречит базальному определению лофофора.
[МГ]: Но если вы возьмете теперь другую пару, то вы тоже найдете у них что-то такое общее, чего нет у третьего.
[ЕТ]: Да, найду.
[МГ]: Это ровно то, что я сказал: когда у нас есть три плохо разрешенных таксона, всегда можно морфологически объединить два против третьего.
[ЕТ]: Поэтому и нужно не только деревья строить, особенно когда их строят люди, которые животных в глаза не видели.
[МГ]: Зачем нужно смотреть животных, чтобы стоить молекулярное дерево?
[ЕТ]: Мне кажется, важно представлять себе, как выглядит животное
[МГ]: Мне тоже кажется, но, с другой стороны, компьютер железный: загрузил в него последовательности, он тебе нарисовал дерево. Какая разница, кто как выглядит? Может быть, наоборот, давление морфологической традиции может быть на какой-то стадии вредным?
[ЕТ]: Есть такое. Скажу честно, я явно под этим давлением нахожусь [смеется].
[МГ]: Хорошо, давайте вернемся. Микроскопы накупили, срезов наделали, на клеточки посмотрели, на строение всего на свете описали, что дальше?
[ЕТ]: Надо посмотреть развитие всего на свете.
[МГ]: В те же микроскопы?
[ЕТ]: В те же микроскопы, но другими методами: экспрессия генов20, мечение бластомеров21… Для этого нужны очень тонкие приборы.
[МГ]: Чем метить?
[ЕТ]: Каким-нибудь флуоресцентным красителем, чтобы выявить потомков этого бластомера.
[МГ]: Вы берете бластомер, закалываете его чем-то светящимся, а потом смотрите, как его потомки распределились по эмбриону?
[ЕТ]: Да.
[МГ]: А еще?
[ЕТ]: Всё.
[МГ]: Я вас все хочу на секвенирование единичных клеток загнать, а вы всё никак не загоняетесь.
[ЕТ]: [Смеется.] Никак не загоняюсь, да. Я пока боюсь этой темы, она для меня совсем новая. Но вообще это очень интересная история, прямо какое-то будущее.
[МГ]: Ожидаете ли вы, что в связи с тем, что молекулярным биологам будет жить хуже, немолекулярным биологам станет лучше?
[ЕТ]: Я думаю, что мы друг без друга не протянем. Надо вместе держаться. У нас тоже есть реактивы, которые станут плохо доступными, если не вообще недоступными. То же оборудование, уже начались сложности с запчастями.

ПРИМЕЧАНИЯ:
1 Филогенетическое древо — дерево, отражающее эволюционные взаимосвязи видов, родов, семейств и других таксономических единиц, произошедших от общего предка.
2 Филогеография — направление географии, которое изучает принципы и процессы (преимущественно эволюционные и экологические), определяющие географическое распределение близкородственных групп организмов.
3 Криптические виды — виды, которые сложно или невозможно отличить друг от друга по морфологическим признакам.
4 Эдификаторы (от лат. aedificator — строитель) — преобладающие в экосистеме организмы, создающие среду обитания для других. Например, коралловые полипы — эдификаторы коралловых рифов, а ель обыкновенная — эдификатор ельника.
5 Билатерии (от лат. Bilatiria), или двусторонне-симметричные, — внеранговый таксон, который включает в себя 99% описанных видов животных. Их общие признаки: сложная нервная система, сквозной кишечник в виде трубки с однонаправленным движением пищи и наличие трех листков в зародыше (экто-, энто- и мезодермы). К Билатериям не относятся лишь четыре типа животных: губки, гребневики, стрекающие и пластинчатые. Остальные 28 типов входят в эту группу.
6 Линяющие (лат. Ecdysozoa) — выделенная Агвинальдо с коллегами группа первичноротых животных, в которую входят членистоногие, нематоды (круглые черви), лобоподы (группа беспозвоночных животных с сегментированным телом и нечленистыми конечностями, в которую, в частности, входят тихоходки), небольшая группа Scalidophora и ряд вымерших таксонов. Линяющих выделили на основе анализа генов рРНК и Hox-генов. Представителям этой группы свойственна линька, а также наличие в покровах особой формы хитина.
7 Аннелиды (лат. Annelida от лат. annelus — кольцо), или кольчатые черви, — многочисленный тип первичноротых животных, один из ключевых общих признаков которых считается сегментированное строение тела. По этому признаку группу аннелид рассматривали родственной членистоногим, однако после пересмотра системы беспозвоночных Агвинальдо с коллегами эти группы животных оказались гораздо более далекими родственниками.
8 Парафилетический таксон — группа организмов, которая включает лишь часть потомков последнего общего предка. Не все подходы современной систематики признают существование парафилитических таксонов: в эволюционной таксономии парафилия рассматривается как частный случай монофилии (происхождения от единого общего предка), в то время как в кладистическом подходе (на котором строится большая часть современной систематики) моно- и парафилия жестко противопоставлены друг другу. В кладистическом подходе только монофилитические группы могут считаться таксонами.
9 Single cell анализ, или транскриптомика отдельных клеток, — недавно появившийся метод, позволяющий изучать транскриптом индивидуальных клетках, то есть какие гены в них работают и насколько активно. Сравнение транскриптомов разных клеток (или даже одних и тех же клеток на разных стадиях развития) позволяет разбираться в молекулярно-генетических основах биологических феноменов. Более подробно о результатах single cell исследований в «Разговорах за жизнь» рассказывали Олег Гусев и Пётр Харченко.
10 Имеется в виду Звенигородская биологическая станция МГУ имени С. Н. Скадовского, где все студенты первого курса биофака МГУ проходят полевую практику.
11 Дорсальный и вентральный (анатомические термины) — соотв. спинной (задний) и брюшной (передний).
12 Ювенильный (биол.) — неполовозрелый.
13 Evo-devo (сокр. от Evolutionary developmental biology) — область биологии, которая изучает родственные связи организмов на основе сравнения стадий их жизненного цикла и эмбрионального развития.
14 Целом — вторичная полость тела животных, представляющая собой заполненное жидкостью пространство между стенкой тела и внутренними органами. От первичной полости тела целом отличается наличием собственной стенки из эпителия. У разных групп животных целом образуется в эмбриогенезе по-разному.
15 Дробление оплодотворенного яйца — самая первая стадия онтогенеза — у разных групп организмов происходит по-разному. По типу симметрии выделяют четыре типа дробления: радиальное (дробление в нескольких плоскостях, но вокруг одной оси), спиральное (дробление вокруг одной оси с разворотом бластомеров по время дробления, которое влечет к «закручиванию» зародыша), билатеральное (имеется плоскость симметрии, из-за чего формируется зеркально симметричный зародыш с левой и правой сторонами) и анархическое (формирование на ранних стадиях несимметричного зародыша, напоминающего цепочку или бесформенную массу клеток).
16 Скребни — тип двухсторонне симметричных беспозвоночных животных, объединяющих червеобразных паразитов.
17 Микрогнатозои — тип двухсторонне симметричных беспозвоночных животных, включающий всего один современный вид Limnognathia maerski, обитающий на моховых подушках в холодных пресных источниках Гренландии.
18 Гнатостомулиды — тип микроскопических (длина самого крупного представителя — 4 мм) двухсторонне симметричных животных, похожих на ресничных плоских червей. От последних гнатостомулид отличает наличие челюстей, по строению близких к челюстям скребней и коловраток.
19 Эктодермальные структуры — структуры, сформировавшиеся в эмбриогенезе из клеток наружного зародышевого листка, или эктодермы. У большинства животных из нее образуется нервная система, кожные покровы и часть пищеварительной системы.
20 Экспрессия генов — процесс, при котором наследственная информация из последовательности нуклеотидов ДНК преобразуется в РНК и (или) белок.
21 Бластомеры — зародышевые клетки, образующиеся на ранних стадиях эмбриогенеза в результате дробления оплодотворенной яйцеклетки (зиготы).